Laboratórna diagnostika má za cieľ určiť v klinických vzorkách pacienta (krv, sérum, moč, stery, výkaly a pod.) prítomnosť infekčného činiteľa. Najcitlivejšou metódou dôkazu na úrovni corpus delicti je priama izolácia mikroorganizmu (baktéria, vírus a pod.) z analyzovaného materiálu, ktorý bol odobratý pacientovi. Žiaľ, k tomuto dôkazu je potrebná bunková kultivácia, t.j. práca s chúlostivejším biologickým materiálom, dostatočne dlhý čas (dni až týždne), a často aj drahá prístrojová technika.
Z praxe vieme, že lekár potrebuje k účinnému zásahu tieto dôležité výsledky čo najskôr. Preto diagnostická prax kladie na životaschopný diagnostický test popri jeho vysokej citlivosti a špecifickosti aj také požiadavky, ako sú rýchlosť, jednoduchosť a ekonomická nenáročnosť. Nie vždy sa darí tieto požiadavky racionálne spojiť. Podmienky diktované praxou podnecujú zrod stále nových diagnostických postupov. Zvlášť výrazne to môžeme pozorovať pri aktuálnej diagnostike vírusových infekcií. Popri priamej izolácii vírusu sa používa napr. dôkaz špecifických vírusových antigénov vyvolávajúcich imunologickú reakciu organizmu (imunofluorescenčný test), dôkaz niektorých vírusových enzýmov (napr. neuraminidázy u chrípkových vírusov, reverznej transkriptázy u retrovírusov), testy založené na špecifických biologických účinkoch vírusov (hematoglutinácia) a pod. Medzi najobľúbenejšie, dobre prepracované testy patria sérologické vyšetrenia založené na princípe dôkazu špecifických protilátok.
Genetický dôkaz mikroorganizmov
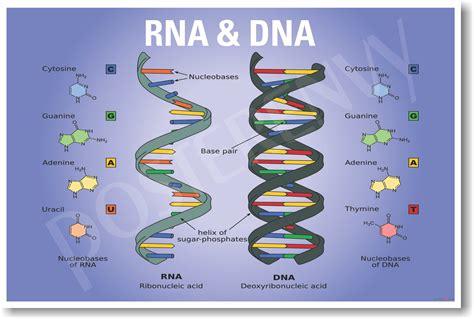
Dnes, v dobe prudkého rozvoja metodík molekulárnej biológie, neprekvapí, že do laboratórnej diagnostiky mikroorganizmov pribúdajú testy práve z tejto oblasti. Všetky sú založené na genetickom dôkaze mikroorganizmu. Mikroorganizmy majú genetický materiál zložený z molekúl DNA alebo RNA (niektoré vírusy). Ak dokážeme vo vyšetrovanej vzorke jeho prítomnosť, určíme pôvodcu infekcie.
Metóda hybridizácie nukleových kyselín
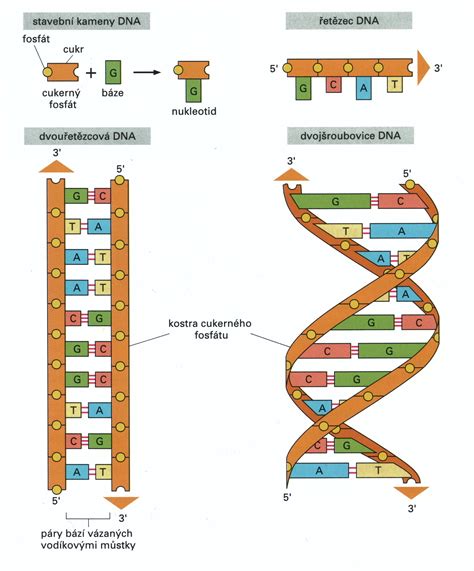
Genetický materiál mikroorganizmov je zložený z reťazca nukleotidov, pričom každý nukleotid obsahuje práve jednu z nukleotidových báz: adenín (A), tymín (T), guanín (G), cytozín (C), v prípade RNA je T nahradený uracilom (U). Nukleotidy vytvárajú jednovláknové alebo dvojvláknové nukleové kyseliny. Dvojvláknové útvary vznikajú špecifickým párovaním báz: A=T, G=C alebo A=U. Iné vzájomné párovanie báz neexistuje.
K dôkazu mikroorganizmu metódou hybridizácie nukleových kyselín je potrebné pripraviť špecifický úsek DNA alebo RNA (sondu), ktorý sledom nukleotidov presne zodpovedá malej (20-30 nukleotidov) alebo väčšej časti (100-2000 nukleotidov) jeho genómu. Ak necháme pri vhodných reakčných podmienkach zreagovať sondu so vzorkou, v ktorej predpokladáme prítomnosť genómu mikroorganizmu, po istom čase (sekundy-hodiny) sa vzájomne zodpovedajúce úseky nukleotidov spoja - zhybridizujú a vytvoria dvojvláknový útvar DNA-DNA, DNA-RNA alebo RNA-RNA. Pretože najlepšie vzájomne hybridizujú jednovláknové úseky nukleových kyselín, dvojvláknové formy genómov mikroorganizmov alebo i sondy sa pred reakciou hybridizácie rozvlákňujú denaturáciou.
Príprava hybridizačných sond
Detekcia mikroorganizmov touto metódou závisí od kvality hybridizačnej sondy. Na detekciu každého typu mikroorganizmu alebo skupiny mikroorganizmov musí byť použitá špecifická hybridizačná sonda. Jej konštrukcia je výsledkom molekulárnobiologických vedomostí o genóme mikroorganizmu, vhodného využitia molekulárnogenetických metodík pri príprave sondy a možností toho-ktorého laboratória. Dnes sa používajú DNA sondy častejšie ako RNA sondy, pretože DNA je stabilnejšia.
Existuje niekoľko spôsobov konštrukcie hybridizačných sond. Jeden z nich využíva princíp, pri ktorom sa z genómu dvojvláknovej DNA (u vírusov sa genóm prepíše do DNA enzýmovým účinkom reverznej transkriptázy) vyštiepi pre daný mikroorganizmus špecifický úsek jednou alebo dvoma reštrikčnými endonukleázami (enzýmy, ktoré štiepia dvojvláknovú DNA v mieste výskytu charakteristického zoskupenia nukleotidov). Pretože pre diagnostické účely potrebujeme väčšie množstvo sondy, využíva sa klonovanie v bakteriálnych bunkách. Vyštiepený úsek sa vsunie do bakteriálneho plazmidu a tento sa vnesie do bakteriálnych buniek, kde sa pomnoží. Po spätnej izolácii takéhoto plazmidu máme k dispozícii dostatok DNA sondy.

Iný spôsob prípravy sondy umožňuje syntéza špecifických fragmentov dlhých 20 - 40 nukleotidov v DNA-syntetizátoroch. V tomto prípade, na rozdiel od predchádzajúcej metódy, je potrebné poznať poradie nukleotidov v genóme mikroorganizmu a vybrať taký úsek, ktorý je pre daný mikrób charakteristický. Pri príprave RNA sond sa využívajú techniky konštrukcie rekombinantných plazmidov a účinky RNA polymeráz.
Zviditeľnenie hybridizačnej reakcie
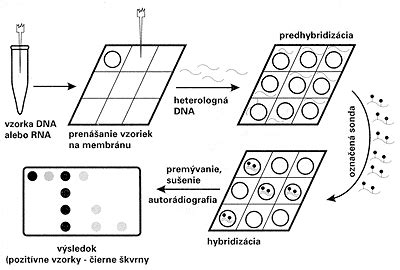
Aj keď sonda zhybridizuje s genetickým materiálom mikroorganizmu, oko experimentátora to nezaregistruje, lebo reakcia prebehne na molekulárnej úrovni. Zviditeľnenie reakcie je možné, až keď sondu označíme. Po rádioaktívnom označení, najčastejšie rádioizotopom 32P, sa hybridizačná reakcia dokáže autorádiografiou, t.j. reakciou rádioaktívneho fosforu (alebo iného rádionuklidu) s citlivou vrstvou filmu. Pretože rádioaktívne značené sondy sú drahé a zdravotne nebezpečné, v menej špecializovaných diagnostických laboratóriách sa nevyužívajú.
Trend jednoznačne smeruje k nerádioaktívnym spôsobom značenia sond. Chemici z rôznych laboratórií odpovedali na túto odbornú výzvu a dnes už máme k dispozícii celú paletu postupov. Najčastejšie sa na nerádioaktívne značenie sond využíva biotín (vitamín H), digoxigenín a pod. Žiaľ, nerádioaktívne sondy sú menej citlivé, ale na druhej strane sú lacnejšie, nepredstavujú zdravotné riziko a sú stabilné niekoľko mesiacov, kým rádioaktívne sondy je potrebné označovať tesne pred ich použitím.
Typy hybridizačných metód
Poznáme množstvo metodických postupov hybridizácie nukleových kyselín. Vzhľadom na metodickú nenáročnosť a rýchlosť sa v diagnostickej praxi často využíva metóda dot-blot hybridizácie s rôznymi technickými modifikáciami. Pri špeciálnych vyšetreniach je možné použiť aj in situ hybridizáciu (určenie lokalizácie DNA alebo RNA priamo v bunke) alebo Southern, resp. Northern hybridizáciu (hybridizácia DNA, resp. RNA po jej štiepení a rozdelení elektroforézou).

Metóda PCR (Polymerázová reťazová reakcia)
Často je detekcia mikroorganizmov sťažená nedostatočným množstvom infekčného agensa v analyzovanej vzorke. Keby však existovala možnosť zvýšiť množstvo genómu mikroorganizmu len chemickým postupom bez potreby kultivácie živých buniek, jeho dôkaz by bol oveľa jednoduchší. Molekulárni biológovia našli kľúč k riešeniu tohto problému prostredníctvom PCR metódy (Polymerase Chain Reaction - polymerázová reťazová reakcia).
Tento nový, neobyčajne jednoduchý metodický postup umožňuje in vitro, t.j. v skúmavke, zmnoženie vybratého úseku dvojvláknovej DNA z nepatrného množstva (teoreticky aj z jednej molekuly) na množstvo rádovo v oblasti nm - mm v priebehu 2 - 4 hodín. K amplifikácii DNA sú potrebné len dva krátke syntetické fragmenty DNA - primery - dlhé 20 - 25 nukleotidov, ktoré majú poradie nukleotidov zodpovedajúce koncom vybratého amplifikovaného úseku DNA. Primery sa spontánne hybridizujú k zodpovedajúcim sekvenciám nukleotidov vo vzorke DNA a po pridaní enzýmu Taq polymerázy sa stanú zárodkom (očkom) pre syntézu nového vlákna DNA.
Amplifikácia (zmnoženie) DNA sa dosahuje len jednoduchou cyklickou zmenou teploty reakčného roztoku, ktorá je dnes automatizovaná termocyklerom, bez potreby pridávania niektorej z reakčných zložiek. Po 20 - 30 cykloch je možné za 2 - 4 hodiny dosiahnuť 106 - 108-násobné zmnoženie vybraného úseku DNA o dĺžke 100 - 2 000 nukleotidov. Keďže po zmnožení máme dostatočné množstvo fragmentu, jeho prítomnosť možno dokázať overením zvolenej fyzikálnej veľkosti (napr. elektroforézou, ktorá trvá len hodinu).
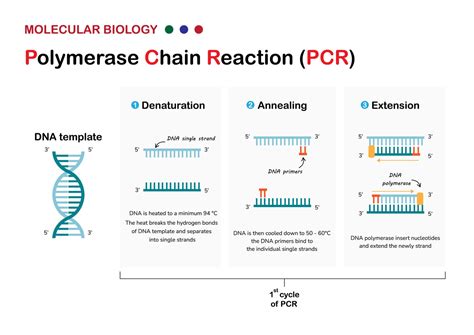
Metodická jednoduchosť a elegantnosť metódy PCR ju priam predurčuje na detekciu mikroorganizmov. V porovnaní s metódou hybridizácie nukleových kyselín je výrazne jednoduchšia, citlivejšia, časove kratšia, nevyžaduje prácu s rádioaktívnym materiálom a možno ju automatizovať. Istým obmedzením je nutnosť poznať nukleotidovú sekvenciu aspoň časti genómu stanoveného mikroorganizmu. Existuje už množstvo prác, ktoré dokumentujú užitočnosť metódy PCR pri vývoji diagnostických testov na herpetické vírusy, HIV, cytomegalovírus, rôzne baktérie a iné mikroorganizmy.
Varianty PCR a izotermická amplifikácia
Pre zvýšenie citlivosti sa niekedy používa metodické usporiadanie dvojitej PCR (nested PCR). Princíp metódy spočíva v tom, že po amplifikácii DNA metódou PCR sa produkt tejto reakcie (napr. DNA o dĺžke 300 bp) opäť amplifikuje v ďalšom teste PCR. Dôjde tým k dvojitej amplifikácii vybratého úseku DNA.
Pri metóde PCR je nutné cyklovanie teploty reakčného prostredia (94 °C - denaturácia vzorky dvojvláknovej DNA, 50 - 60 °C - väzba primerov, 72 °C - optimálna teplota pre Taq-polymerázu pri syntéze nových vlákien DNA). Existuje však postup, pri ktorom amplifikácia DNA prebehne pri konštantnej izbovej teplote, napríklad pomocou amplifikácie sondy s enzýmom QBeta-replikáza. Špecifická sonda sa nahybridizuje k stanovenej vzorke nukleovej kyseliny a nenaviazaná časť sondy sa odstráni. V ďalšom kroku sa uvoľní nahybridizovaná sonda zo vzorky, ale keďže je v malom množstve, ťažko sa dokazuje. Preto je potrebné ju namnožiť. Využíva sa pritom enzým QBeta-replikáza, ktorý je schopný už za 30 minút pri stálej teplote 20 °C amplifikovať sondu viac ako biliónkrát (napr. 0,14 fg nukleovej kyseliny zmnoží na 130 ng).
Výhody molekulárnogenetických testov
Molekulárnogenetické testy ponúkajú niekoľko významných výhod. Ako prvú výhodu uveďme možnosť diagnostiky infekčných chorôb na základe dôkazu genómu infekčného činiteľa bez potreby kultivácie buniek, pričom je zachovaná vysoká citlivosť a špecifickosť. Z tohto základného faktu sa odvíja ďalšia prednosť: infekčný proces je možné objaviť už v rannom štádiu, kedy môže byť lekársky zásah veľmi účinný.
Výraznou výhodou takýchto testov je možnosť diagnostiky latentných stavov infekcií, kedy sú mikroorganizmy v pokojovom stave, takže mnohé klasické diagnostické testy sú nepoužiteľné. Využitie týchto metodík možno očakávať napr. u vírusov, ktoré sa ťažko množia na bunkových kultúrach alebo u mikroorganizmov potláčajúcich imunitnú odpoveď hostiteľa. Domnievame sa, že molekulárnogenetické postupy napriek svojmu prudkému rozvoju nevytlačia úplne doterajšie, mnohokrát osvedčené testy mikroorganizmov.
Princípy izolácie nukleových kyselín
Izolácia nukleových kyselín je prvým krokom v mnohých experimentoch molekulárnej biológie. Ideálne je potrebné získať cieľovú nukleovú kyselinu (DNA alebo RNA) v dostatočnom množstve a kvalite, to je bez prímesí „nadbytočných” látok, tzv. kontaminantov. Nukleové kyseliny je možné izolovať z rôznych zdrojov. Pre úspešnú izoláciu je nevyhnutné izolovať čistú, vysokomolekulárnu DNA, hoci pre niektoré aplikácie, ako napríklad analýza mutácií, postačujúca je fragmentovaná DNA s dĺžkou fragmentov 70-150 kb.
Výber metódy izolácie záleží od spôsobu, akým sa bude nukleová kyselina v ďalšom postupe analyzovať. Napr. pri použití metódy PCR (polymerázová reťazová reakcia) postačujú malé množstvá DNA, ktorej čistota a integrita nie sú vo väčšine prípadov kritické. Na druhej strane, pri klonovaní, enzymatickom opracovaní DNA reštrikčnými endonukleázami alebo pri sekvenovaní sú potrebné relatívne väčšie množstvá DNA s vyššou čistotou.
Identifikácia bakteriálnej DNA sa skladá z niekoľkých krokov: lyze buniek, izolácia a purifikácia DNA, precipitácia ethanolom a identifikácia bakteriálneho kmeňa pomocou PCR, prípadne aj iných molekulárno biologických metód. Každý krok musí byť optimalizovaný.
Lyza buniek: Prvý krok izolácie DNA
Aby došlo k uvoľneniu nukleovej kyseliny do vodného roztoku, je potrebné rozrušiť (dezintegrovať) bunkové steny a biomembrány. V prípade, že sa bunková kultúra pestuje v tekutom médiu, je prvým krokom izolácie oddelenie buniek od média centrifugáciou.
Vo všeobecnosti sa ťažšie izolujú nukleové kyseliny z rastlinných buniek kvôli prítomnosti kompaktnej celulózovej bunkovej stene a chemických zlúčenín obsiahnutých vo vakuolárnej šťave. Na uvoľnenie obsahu rastlinných buniek obalených bunkovou stenou sa používa v prvom kroku mechanické ako aj enzymatické opracovanie (celulázy).
V prípade niektorých baktérií, ktoré majú veľmi hrubú bunkovú stenu alebo tvoria veľa sekundárnych metabolitov, je výhodné izolovať DNA z buniek v logaritmickej fáze rastu, t.j. pri ich aktívnom hromadnom delení. Na lýzu bakteriálnych buniek sa najčastejšie používa enzymatické opracovanie bunkovej steny lyzozýmom, ktorý hydrolyzuje glykozidické väzby v peptidoglykánoch. Tento enzým sa prirodzene vyskytuje vo vaječnom bielku a v slzách. Druhým krokom je rozrušenie cytoplazmatickej membrány bakteriálnej bunky pôsobením ionogénnych detergentov (detergentov tvoriacich ióny), napr. SDS − dodecylsulfát sodný. Nevzácne je treba okrem chemickej lýzy použiť aj mechanickú lýzu bakteriálnych buniek pomocou ultrasonikátora, ktorý na disrupciu buniek využíva ultrazvukové vlny.
V prípade izolácie nukleových kyselín zo živočíšnych tkanív a celých orgánov (myšacia slezina, pečeň, mozog) je potrebná homogenizácia vstupného materiálu. K tomuto účelu je možné použiť mechanické rozdrvenie v homogenizátore alebo rozrušenie bunkovej hmoty zmrazením v tekutom dusíku. Živočíšne bunky sú obalené len cytoplazmatickou membránou, a preto je možné dosiahnuť ich lýzu jemnejšími prostriedkami, napr. pôsobením slabých neionogénnych detergentov (napr. Triton X-100) v kombinácii s proteolytickými enzýmami (napr. proteináza K).
K rozpadu bunkových štruktúr prispieva degradácia proteínov pomocou enzýmov proteáz, napr. proteinázy K. Dôležitým faktorom je aj prítomnosť chelatačných činidiel (napr. EDTA − kyselina etyléndiamíntetraoctová), ktoré z roztoku „vychytávajú” dvojmocné katióny (Mg²⁺, Ca²⁺), čím inaktivujú bunkové DNázy. Neprítomnosť dvojmocných katiónov zároveň destabilizuje vonkajšiu bakteriálnu membránu. Pri lýze sa uvoľní celý obsah buniek do roztoku, pričom vznikne komplexná zmes obsahujúca zvyšky bunkových stien a bunkových membrán, proteíny, nukleové kyseliny, polysacharidy a rôzne nízkomolekulové zložky (obzvlášť dôležité pri izolácii z rastlinných buniek kvôli prítomnosti chemických zlúčenín vakuolárnej šťavy). Aby nedochádzalo k degradácii izolovanej nukleovej kyseliny je potrebné použiť čo najmiernejšie podmienky lýzy, t.j. uskutočňovať ju v tlmivom roztoku a v chlade.
Purifikácia a koncentrácia DNA
Princípom purifikácie akéhokoľvek materiálu z buniek je cieľom zbaviť sa zložiek, ktoré pri ďalšej práci nepotrebujeme. Pri izolácii nukleových kyselín je dôležité odstrániť zo zmesi proteíny, medzi ktoré patria aj bunkové nukleázy, ktoré môžu degradovať izolovanú nukleovú kyselinu, ako aj proteíny viažúce sa na DNA, ktoré svojou prítomnosťou môžu rušiť následné analýzy (napr. štiepenie reštrikčnými endonukleázami).
Odstránenie proteínov a RNA
Na odstránenie proteínov z preparátov nukleových kyselín sa často používa extrakcia organickými rozpúšťadlami (fenol, chloroform = trichlórmetán) v pomere 24 : 1 (fenol/chloroform-izoamylalkohol). Tieto látky sú s vodou nemiešateľné, a preto sa po ich pridaní k vodnému roztoku nukleových kyselín vytvoria dve fázy. Pri premiešaní takejto zmesi dochádza na fázovom rozhraní k denaturácii proteínov. Ak sa roztok nukleových kyselín extrahuje fenolom s neutrálnym pH, nukleové kyseliny zostávajú po extrakcii vo vodnej fáze. Po fenolovej extrakcii je dôležité zbaviť roztoky nukleových kyselín zvyškov fenolu, pretože fenol môže inhibovať následné enzymatické reakcie (napr. reštrikčné štiepenie). Jeho veľkou nevýhodou je vysoká toxicita. Izoamylalkohol zas zlepšuje separáciu vrstiev pri centrifugácii a udržuje ich stabilitu.
Na odstránenie RNA z preparátov DNA sa používa pankreatická RNáza. Tento enzým je veľmi stabilný a zároveň nevyžaduje pre svoju aktivitu dvojmocné ióny, čo neplatí napr. pre DNázy.
Alkoholové zrážanie a elúcia DNA
Na prečistenie nukleových kyselín od nízkomolekulových látok (soli, zvyšky fenolu) a na skoncentrovanie vzorky sa používa alkoholové zrážanie (najčastejšie čistým, bezvodým etanolom alebo izopropanolom). Zrážanie sa robí v prítomnosti jednomocných iónov (napr. 0,1 M octan sodný) a pri nízkej teplote (-20 °C). V týchto podmienkach je výťažok kvantitatívny aj pri nízkej koncentrácii DNA. Na druhej strane, zrážanie pri laboratórnej teplote (+20 °C) je výhodné, ak chceme vyzrážať DNA bez súčasného vyzrážania („oživenia”) kontaminujúcej RNA.
Vyzrážaná nukleová kyselina sa nazýva precipitát. Precipitát DNA sa od roztoku oddelí vysokootáčkovou centrifugáciou. Premytím 70 % etanolom sa odstránia zvyšky solí prítomné v precipitáte a precipitát sa vysuší tak, aby sa odparili zvyšky etanolu, ale zároveň aby DNA zostala hydratovaná. Molekula DNA v kyslom prostredí môže spontánne degradovať, a preto sa DNA rozpúšťa v slabo zásaditom tlmivom roztoku s prídavkom EDTA (tzv. roztok TE).
Špecifické metódy izolácie DNA
Zdrojom nukleových kyselín môžu byť aj subcelulárne častice - organely alebo vírusy, ktoré získame po izolácii centrifugáciou v hustotnom gradiente. Ten pozostáva zo starostlivo namiešaného roztoku (najčastejšie sacharózy) s najväčšou koncentráciou naspodu, na ktorý postupne nalievame roztoky s nižšou koncentráciou, pričom pri pozornej príprave sa tieto roztoky nezmiešajú ale ostanú ako oddelené fázy. Pri centrifugácii bunkového materiálu v takomto hustotnom gradiente dochádza k zachytávaniu rôzne ťažkých sedimentov na určitom medzistupni/fáze.
Metóda alkalickej lýzy sa používa pri izolácii plazmidov z bakteriálnych buniek za súčasného odstránenia chromozomálnej DNA. Bakteriálne bunky sú lyzované v alkalickom prostredí (0,2 M NaOH) v prítomnosti detergentu (1 % SDS). Chromozomálna DNA je pri lýze fragmentovaná na lineárne molekuly, zatiaľ čo plazmidová DNA, ktorá má podstatne menšiu veľkosť, si zachová cirkulárnu formu. Pri pH okolo 12, ktoré je v roztoku, dochádza k denaturácii DNA, t.j. vodíkové väzby medzi bázami sa uvoľnia a vznikajú jednovláknové molekuly. Po neutralizácii roztoku (octanom draselným) kruhová plazmidová DNA renaturuje do pôvodnej dvojvláknovej formy, ale lineárne molekuly chromozomálnej DNA nie sú schopné tak rýchlo renaturovať, a preto agregujú s inými zložkami lyzátu (bunkové steny a membrány, proteíny, SDS).
DNA môže byť purifikovaná zo zmesi makromolekúl ultracentrifugáciou za pomoci chloridu cézneho (CsCl) a etídium bromidu. DNA sa skoncentruje do úzkeho prúžku v strede gradiantu, pričom lineárna a cirkulárna (plazmidová) DNA majú inú hustotu, a preto je možné ich touto metódou navzájom oddeliť. Po centrifugácii sa vyberú frakcie obsahujúce DNA, etídium bromid sa odstráni extrakciou n-butanolom a DNA sa z roztoku získa etanolovou precipitáciou. Výsledkom tejto techniky je vysokomolekulárna DNA s vysokou čistotou.
Izolácia RNA
Pri izolácii RNA z akéhokoľvek biologického materiálu je hlavným problémom jej nestabilita. Molekula RNA podlieha degradácii oveľa ľahšie ako DNA a zároveň ribonukleázy sú vo všeobecnosti veľmi stabilné a všadeprítomné enzýmy, ktoré nepotrebujú k svojej aktivite ako kofaktory dvojmocné ióny, na rozdiel od DNáz.
Vedeli ste, že...? RNáza sa skutočne vyskytuje všade. Vylučuje sa aj potnými žľazami na končekoch prstov, takže práca bez rukavíc je absolútne neprípustná.
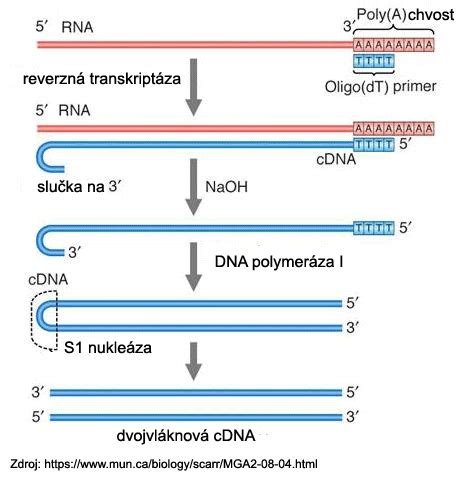
Metódy izolácie a purifikácie RNA sú založené na rýchlej lýze buniek, ktorá efektívne neutralizuje prítomné nukleázy. Kontaminujúce bielkoviny sa odstraňujú fenolovou extrakciou v kyslom prostredí alebo na silikagélových kolónkach. Zvyšky chromozomálnej DNA je možné rozložiť pôsobením DNázy. Purifikovaná RNA sa využíva na prípravu komplementárnej DNA (cDNA), northernovu hybridizáciu, RT-PCR, alebo na in vitro transláciu.
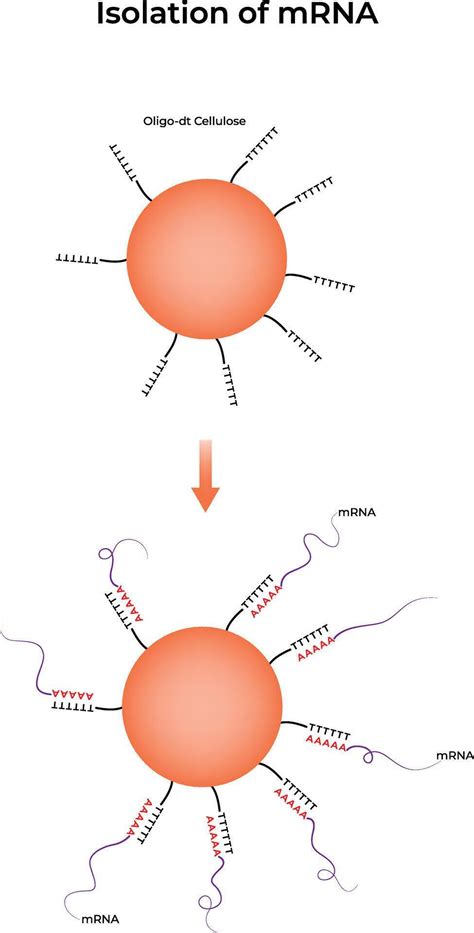
Pri mnohých aplikáciách je výhodné použiť iba frakciu mRNA. V princípe, izolácia mRNA spočíva vo využití prirodzenej enzymatickej posttranskripčnej modifikácie eukaryotickej mRNA, a to je pridávanie poly(A)-chvostíka na 3'-koniec molekuly mRNA za pomoci enzýmu poly(A)-polymerázy. Oligo-dT nosiče sú jednoduché plôšky, na ktorých sú fixované jednovláknové molekuly so sekvenciou poly(T), a teda sú komplementárne k jednovláknovým mRNA s poly(A) koncami. Ak sa roztok s prítomnými mRNA naleje na tieto nosiče, v prostredí s vyššou iónovou silou (pridanie solí) dochádza k hybridizácii, a teda zachytávaniu mRNA na týchto nosičoch.
Izolácia DNA pomocou magnetických nosičov
Nukleové kyseliny možno z buniek s výhodou izolovať pomocou magnetických nosičov. Táto špecifická a jednoduchá technika pre izoláciu DNA sa opiera o elektrostatickú interakciu DNA s povrchom častíc. Magnetické častice, často magnetit (Fe3O4) a maghemit (γ-Fe2O3), môžu byť povrchovo upravené oxidmi (oxid kremičitý, oxid hlinitý).
Molekuly DNA sú na povrch magnetického nosiča naviazané na základe elektrostatických interakcií a posléze sú eluované do pufra. Použitie magnetických nanočastíc pre aplikácie v biomedicíne a biosnímaní je stále rozsiahlejšie.

Optimalizácia izolácie bakteriálnej DNA s magnetickými nosičmi
Cieľom práce je optimalizácia jednotlivých krokov identifikácie bakteriálnej DNA: bunková lyza, izolácia DNA, charakterizácia pevných magnetických nosičov funkcionalizovaných aminoskupinami pre izoláciu nukleových kyselín. Každý krok musí byť optimalizovaný.
Proces izolácie bakteriálnej DNA zahŕňa inkubáciu lyzátu so 100 μl magnetických častíc a 5 M NaCl. Zmes sa premieša a potom sa postaví na magnetický stojanček po dobu dvoch minút, čo umožní adsorptívny prenos. Supernatant sa odstráni, pričom sa dbá na to, aby neboli nasaté častice. Následne sa promývajú sedimentované častice Tris-HCl.
Prítomnosť DNA v eluáte bude overená pomocou gélovej agarosovej elektroforézy a množstvo eluovanej DNA bude stanovené spektrofotometricky. Kvalita izolovanej DNA bude overená jej amplifikáciou s použitím polymerázovej reťazovej reakcie (PCR).
Vplyv eluovacieho pufra na výťažnosť DNA
Bolo zistené, že pH eluovacieho roztoku Tris-EDTA je optimalizované pre vyššiu výťažnosť izolácie DNA. Opakovateľnosť však môže byť zhoršená.
Pre porovnanie boli testované rôzne eluovacie roztoky, vrátane Tris-EDTA o pH 6,5 a Tris-HCl a Tris-EDTA o pH 9. Najvyššia výťažnosť (100 %) bola stanovená u roztoku Tris-EDTA o pH 9. V prípade rovnakého roztoku o pH 6,5 bola výťažnosť 36,9 %.
Pri použití H2O ako eluovacieho roztoku bola RSD 18,1 %. Výťažnosť za týchto podmienok bola nižšia ako v predchádzajúcom prípade (34-37 %). Pre fosfátový pufer I bola RSD 1,2 %, no výťažnosť bola za týchto podmienok nízka (23,5 %).
| Elučný roztok | pH | Výťažnosť DNA (%) | RSD (%) |
|---|---|---|---|
| Tris-EDTA | 9,0 | 100 | N/A |
| Tris-EDTA | 6,5 | 36,9 | N/A |
| H2O | N/A | 34-37 | 18,1 |
| Fosfátový pufer I | N/A | 23,5 | 1,2 |
Tieto výsledky ukazujú, že výber eluovacieho roztoku je kritický pre dosiahnutie maximálnej výťažnosti DNA.
Aplikácie molekulárnogenetických metód a ďalšie štúdie
Okrem izolácie DNA pre diagnostické účely, sa práca zameriava aj na štúdium sekundárnych štruktúr nukleových kyselín - krížových štruktúr a kvadruplexov. Tieto štruktúry sa podieľajú na regulácii bunkových procesov a ich výskyt je spájaný s vývojom onkologických a neurodegeneratívnych ochorení.
Bola vykonaná analýza genómu in silico u mikroorganizmov významných v potravinárskom priemysle. Sekvencie genómov mikroorganizmov boli získané z databázy NCBI (National Center for Biotechnology). Pre analýzu bol použitý softvér Palindrome Analyser a G4 Hunter.
Molekulárno-genetické testy majú široké uplatnenie v diagnostike rôznych ochorení, vrátane identifikácie rakoviny ústnej dutiny a iných ochorení pomocou analýzy mutácií.
tags: #izolacia #dna #mikroorganizmov